
| |
| Volume 16, Number 1 | June 9, 2026 |
This issue, I share with you a recent conceptual breakthrough. I regard it as a birthday gift.
I normally wake up around 6:30 a.m., but one morning, a few months back, I woke up at 5:00. My nose had started to run, and it woke me up. Helen and I had gone out to eat at a restaurant a few days previously. It seems I had picked up a cold while we were there.
I have a recliner with a blanket in my home office for such occasions, to keep from ruining the rest of Helen's night's sleep. My nose would give me no peace, even while sitting somewhat upright under the blanket in my recliner. So I gave up trying to get back to sleep and started working, in my head, on a problem I have worked on, off and on, for years. The problem is how exactly vitamin MePiA manages to quench free radicals in the mitochondria without being lost to chemical reaction. Biblical life span data show that MePiA has an incredibly long (centuries) biological half-life.[1] So somehow MePiA must get recycled. But how?
I had started to tackle this problem again the previous day, which is why it was on my mind when I woke at 5:00. I had mentioned it to Helen at supper and shared that it was a very difficult problem which I had invested time in over the years with some modest gains, but without so far managing to get all the puzzle pieces to fit properly together. The research had now come to the place where this problem really needed to be solved.
Helen said, "Don't worry. You'll have it solved before another 24 hours has passed."
"Most improbable," I thought to myself. But it is a wonderful thing to be married to such a woman. Research hogs time, leaving little over to "do life." And it hogs money. We have lived much of our married lives below the poverty line. And research is often emotionally draining. Ups and downs and twists and turns are inevitable for any real research program. Helen has lived with the ecstasies and agonies of my research for decades. Yet here she was, as usual, nothing but wind in my sails.
As I sat in my recliner, staring into the darkness, concentrating, following the logic, bringing to the fore the critical questions, I suddenly cleared a conceptual hurdle, and the pieces began to fall into place.
How did Helen know?
The answer to this longstanding MePiA mystery is now recorded in my current lab book: Life Span Research Laboratory Notebook 34, page 91. It is recorded as a continuation from the last previous entry on the same topic: Life Span Research Laboratory Notebook 29, page 10. The date of the previous entry is November 25, 2016. Only when I had written the date on the page of the current entry, February 18, 2026, did I remember that it was my birthday.
MePiA's role is all about free radicals. That much I have known for a long time.[2] But I have thought of MePiA as soaking up free radicals—specifically, soaking up reactive oxygen species (ROS)—converting chemically to MePA in the process (Figure 1).[3]
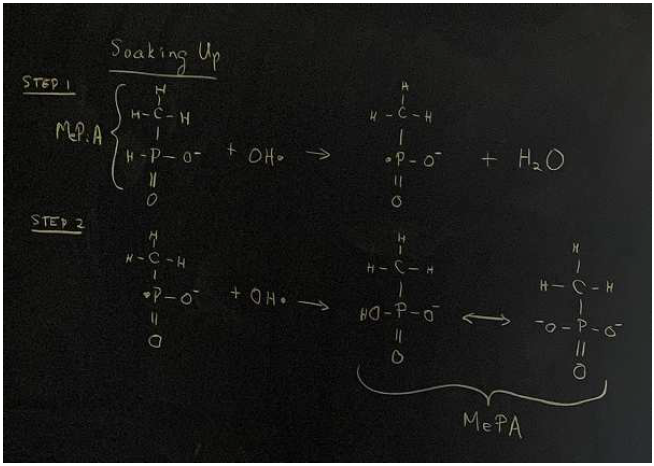 |
The spontaneous conversion of MePiA to MePA is easily observed in the laboratory. Measure the amount of MePiA and MePA in an aqueous solution. Cap the solution and let it sit at room temperature. Cosmic rays will produce free hydroxyl radicals from water molecules in the solution. After a month, measure the solution again. You will observe that MePiA has decreased and MePA has increased. This observation, which I have made numerous times for various reasons, lies at the root of my conception of MePiA as soaking up free radicals.
This conception leads to the problem of how the body manages to recycle MePA back to MePiA so that MePiA is conserved and not lost.
After much digging into how MePA might be converted back to MePiA, it appeared to be not only a difficult thing to do but also likely an impossible thing to do. To this personal assessment, new-on-the-scene AI has now added its assurance.
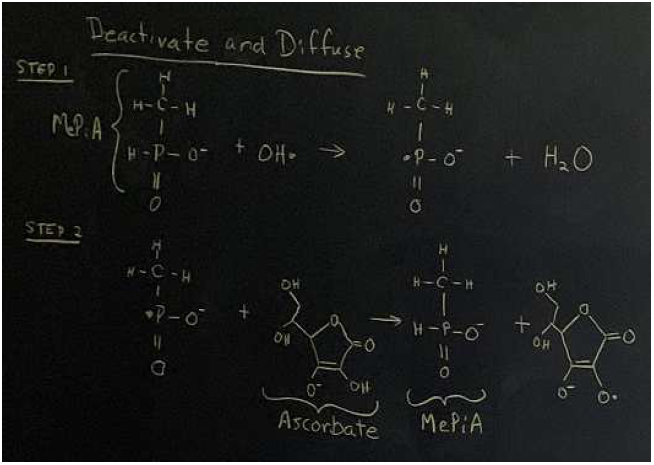
My conception of MePiA soaking up free radicals turns out to be wrong for the mitochondrial matrix. It is wrong because, in the matrix, the concentration of antioxidants is vastly greater than the concentration of free radicals, so the probability of a given MePiA molecule interacting with two free radicals in a row is miniscule. The present article proposes my new conception that, within the mitochondrial matrix, MePiA merely deactivates and diffuses free radicals (Figure 2). In this conception, MePiA acts only as the front line of defense. It does not do the whole job by itself alone. It is supported by existing, well-known second-line defense mechanisms involving antioxidant molecules, such as vitamin C and glutathione, which recycle it back to MePiA.
 |
The hydrogen atom (H) is well known as the simplest example of an atom. Not so well known is the fact that it is also the simplest example of a free radical. It is made up of a positively charged central proton with a single, negatively charged electron orbiting the proton. The fact that the hydrogen atom has an orbital electron which is unpaired is what makes it a free radical. The free radical character of hydrogen can be made explicit by representing its unpaired, orbital electron as a lone dot, H·.
In the world of atoms and molecules, single electrons don't like to remain single. Electrons strongly prefer to be paired.
In quantum mechanics, we talk about electrons having a spin, and we say that the spin can be in only one of two states: up or down. Electrons prefer to be paired, one with spin up and the other with spin down. That's the way God has made the world to be.
Now, if you purchase a container of hydrogen gas, you will not get a whole lot of individual hydrogen atoms in the container. You will get a whole lot of paired hydrogen atoms, H-H (usually written as H2), in the container. The hydrogen atoms have all paired up. In each pair, the two hydrogen atoms are stuck (or bonded) together. As H is the simplest atom, so H2 is the simplest molecule. The container will be filled with hydrogen molecules, H2, not hydrogen atoms, H, because H is H·, and the single electrons in all of the hydrogen free radicals want to pair up. So the electron of one hydrogen atom will pair with the electron of another hydrogen atom. This causes the two hydrogen atoms to bond together into a single hydrogen molecule.
The bond is often denoted as a dash, H-H, but the bond is actually made up of two electrons, so the character of the bond may be made more explicit by representing it as a pair of dots, H:H.
Now you can see that a hydrogen molecule, H:H, may be thought of as two free hydrogen radicals, H·, which have become two bound radicals, H:H.
Bound radicals are happy to stay that way and are thus relatively unreactive chemically. Free radicals are not happy to stay that way and are very reactive chemically. And all of this leads to the simple rule that a free radical may be tamed (quenched) by binding it to another radical. This, in fact, is what MePiA does—what MePiA is very good at doing—as I will show shortly below.
The hydrogen radical is very important biologically, though it mainly fills its biological role as a bound radical, not as a free radical. Important biological free radicals are mostly of the oxygen atom (O) type. This family of free radicals is called ROS (Reactive Oxygen Species).
Oxygen, like hydrogen, occurs in nature as a molecule (O2), not a lone atom (O). But it does so for a different reason—another one of God's rules for quantum mechanics which we need not go into here.
In contrast to hydrogen, O is not a radical. While H has only 1 proton, and hence 1 electron, O has 8 protons in its nucleus, and hence 8 orbital electrons. This yields 4 pairs of electrons orbiting the O nucleus. As there is no unpaired electron with O, it is not a free radical. But it can easily give rise to free radicals.
There are two derivatives of O—two ROS—which are particularly of interest in the present context. These are 1) superoxide, O2-·, and 2) hydroxyl radical, OH·. Both are potentially destructive of mitochondria because of their chemical reactivity, but of the two, OH· is the more reactive and therefore the more important to control.
Because OH· is more potentially destructive than O2-·, it comes as a surprise to learn that while O2-· has a special enzyme, called superoxide dismutase (SOD), dedicated to quenching it, OH· has no analogous special component dedicated to quenching it.
I propose that this apparent omission is a smoking gun. I will come back to this smoking gun in a moment, after a brief further discussion of O2-·.
Superoxide, O2-·, is an inevitable side product of oxidative phosphorylation (OXPHOS). Meanwhile, OXPHOS is essential for the provision of adequate energy to cells. OXPHOS takes place in mitochondria. It is pretty much what mitochondria are all about. OXPHOS entails a flow of electrons. Whenever an electron "leaks" from this flow and attaches to an O2 molecule, the result is superoxide.
The O2 molecule is not a free radical. But O2 has the property that it readily accepts another electron, so it readily accepts electrons which have leaked from OXPHOS. When this happens, it does two things: 1) it turns the oxygen molecule into a negatively charged oxygen molecule, O2-, because the added electron adds its negative charge to the O2 molecule; and 2) it simultaneously turns the oxygen molecule into a free radical molecule, O2-·, because the added electron is not paired. This negatively charged, free radical oxygen molecule, O2-·, is superoxide.
Because O2-· is a free radical, and hence chemically reactive, it has potential to do a great deal of harm to mitochondria. Mitochondria are made of biomolecules. Superoxide can break biomolecules in pieces or otherwise convert them into different molecules no longer suited to their original biomolecular jobs.
As mentioned above, there is a special enzyme, SOD, which quenches O2-·. It does so by catalyzing conversion of superoxide into hydrogen peroxide, HOOH (also designated H2O2) and O2. I will not go into how it does this except to mention that it is pretty slick—hats off to the Creator once again.
You can see immediately that HOOH is not a free radical because there is no free electron. Hydrogen peroxide is made up of two oxygen atoms, which, as mentioned above, are not free radicals, plus two H radicals, which, when combined in the HOOH molecule, yields only another set of paired electrons. But HOOH can react with iron ions present within the matrix to yield OH·. This is how OH· comes to be present in the mitochondrial matrix.
So SOD does not entirely put an end to the free radical problem created by O2-·. In fact, in one way, it makes the problem worse. While the production of O2-· is localized near the oxidative phosphorylation apparatus on the inner cell wall, the production of OH· is not localized. Hydrogen peroxide is a small, water-like molecule. It can diffuse anywhere and everywhere throughout the matrix. This means that OH· is able to be generated anywhere and everywhere throughout the matrix, damaging anything and everything within the mitochondrial biological machine. Most importantly, it will have access to the mitochondrial DNA (mtDNA), damaging and mutating it.
In this context, omission of a front line of defense against OH· seems most unlikely to be a design feature. Indeed, it strikes one immediately as a design flaw. How can there be no component deliberately designed to deactivate OH·, the worst of the ROS lot with potential to do such widespread and critical damage within the mitochondrial matrix? I contend that this apparent omission is a smoking gun having the fingerprints of dietary deficiency of MePiA all over it.
I propose that MePiA is meant to function as the front-line antioxidant for hydroxyl radicals. That is, it is meant to deactivate hydroxyl radicals, quenching them to harmless water molecules. In the process, MePiA itself becomes a free radical, but this free radical is not nearly as reactive as the hydroxyl radical. It is quenched by subsequent, well-known antioxidant mechanisms within the mitochondrial matrix, recycling it back to MePiA.
In short, I propose that MePiA is to OH· what SOD is to O2-·. The fact that MePiA is absent from the environment today, and thus absent from human diets today, is why it is absent from the mitochondrial matrix today. This is all a consequence of the Flood of Noah, a real-world historical, global geophysical catastrophe with real-world consequences, as I have shown repeatedly.[4] This loss of MePiA from the mitochondria is the fundamental physiological explanation of the second of the two aging diseases created by the Flood: 1) MePA deficiency disease and 2) MePiA deficiency disease. MePiA deficiency disease results in MePiA missing from the mitochondrial matrix, which removes from the mitochondria their natural front line of defense against the biomolecular depredations of OH·. The result is accumulating mitochondrial damage and dysfunction, culminating ultimately in sickness and death of the organism.
MePiA is well suited to the job of front-line defense against OH· for several reasons.
First, MePiA is an antioxidant. This is essential. The entire purpose in this case is to quench the free radical OH·, and quenching free radicals is what antioxidants do.
A feature of many antioxidants is their ability to give up a hydrogen atom easily. The strategy here is really very simple. Hydrogen, we now know, is a radical. If a hydrogen radical is given up to some free radical, the two radicals will bind, quenching the free radical.
In the case of MePiA, the H which is bonded to the phosphorus atom (P) is easily given up. The reason for this begins with the fact that MePiA is an acid. MePiA is methylphosphinic acid. Acids readily give up protons. At physiological pH, nearly all MePiA exists as the negative ion, methyl phosphinate (MePiA-), having given up a proton. But giving up a proton does not quench any free radicals. For that job, the full hydrogen atom is needed, the proton plus an electron. But the fact that MePiA, in the matrix, has already given up a proton makes it especially susceptible to subsequently surrendering a whole hydrogen atom.
Focus on the central phosphorous atom in STEP 1 of Figure 1 or Figure 2. It is bonded with a hydrogen atom. If this phosphorus atom is made rich in negative charge, Coulomb repulsion between the electrons will encourage loss of an electron. MePiA- has an obvious excess of negative charge relative to MePiA. It has an O atom which has lost a chemically bonded proton, decreasing the overall positive charge of the molecule and thereby increasing the negative charge density of the molecule. But this excess negative charge density merely adds to an already high negative charge density for the phosphorus atom. This phosphorus atom also has a double-bonded O attached to it, giving it four more electrons to deal with, and it has an electron-donating methyl group (CH3) attached to it, feeding it yet more electrons. This high density of electrons local to the phosphorous atom makes it relatively easy for the hydroxyl radical to take the hydrogen atom from the phosphorous atom, proton plus electron.
This is how MePiA- quenches OH·. It readily gives up a hydrogen atom to OH·. This pairs the two dots (the two lone electrons) deactivating them, and it adds a second H to the OH, yielding H2O—water. In this way, MePiA- converts the dreadful OH· to a harmless water molecule.
Not every molecule is an antioxidant. The vast majority of molecules are not antioxidants. And not every antioxidant is suited to front-line defense against OH·. Most are useless against mitochondrial OH·. Hydroxyl radicals are simply too reactive for them to handle, but MePiA- is a small, robust molecule, able to surrender a hydrogen atom to OH· without being destroyed in the process. This property is essential, of course, for later recycling back to MePiA-, and this is essential to explaining the extraordinary biological half-life of MePiA.
Now here is another way that MePiA is well suited to the job of front-line defense against OH·. The free radical which it becomes when H is abstracted from it by OH· is a relatively tame free radical. The presence of the two oxygen atoms bonded to the P radical center effectively spreads out the unpaired electron, reducing its reactivity. This gives time for the abstracted H radical to be replaced by second-line defense mechanisms before this free radical inflicts damage on any surrounding biomolecule.
The idea that MePiA soaks up free radicals in the mitochondrial matrix by reacting with two hydroxyl radicals in two steps, converting to MePA in the process, is mistaken. Rather, one should think of MePiA as reacting with a single hydroxyl radical in a first step only, as a front-line defense, quenching the hydroxyl radical to a water molecule by giving up a hydrogen radical to it. This converts MePiA itself into a free radical, but this new free radical is much less reactive than the hydroxyl radical, enabling it to be quenched back to MePiA by second-line antioxidants such as vitamin C and glutathione.
This new view has large potential consequences, entirely interrupting the direction of research outlined last issue.[5] It calls into question a key assumption. I have assumed, from very early on, that the body pool of MePiA was tiny—micrograms. This new view raises the possibility that it may be large—grams. If it is large, then it is possible that the recommended daily intake (RDI) of MePiA needs to be much larger than previously thought.
I have now spent several months reexamining this RDI via measurements of urinary excretion of MePiA versus intake amount. I still do not know the size of the body pool of MePiA, but it presently appears that the RDI for MePiA should be increased.
I expect to share more of all of this, together with final results of this urinary excretion experimental program, next issue. ◇
The Biblical Chronologist is written and edited by Gerald E. Aardsma, a Ph.D. scientist (nuclear physics) with special background in radioisotopic dating methods such as radiocarbon. The Biblical Chronologist has a fourfold purpose:
The Biblical Chronologist (ISSN 1081-762X) is published by: Aardsma Research & Publishing Copyright © 2026 by Aardsma Research & Publishing. |
^ Gerald E. Aardsma, Aging: Cause and Cure, 3rd ed. (Loda, IL: Aardsma Research and Publishing, 2023), 212. www.BiblicalChronologist.org.
^ Gerald E. Aardsma, Addendum to Aging: Cause and Cure (Loda, IL: Aardsma Research and Publishing, July 26, 2019), 10. www.BiblicalChronologist.org.
^ Gerald E. Aardsma, Aging: Cause and Cure, 3rd ed. (Loda, IL: Aardsma Research and Publishing, 2023), 131. www.BiblicalChronologist.org.
^ See, for example, Gerald E. Aardsma, Noah's Flood Happened 3520 B.C. (Loda, IL: Aardsma Research and Publishing, 2015). www.BiblicalChronologist.org.
^ Gerald E. Aardsma, "The End of Modern Human Aging Appears Imminent," The Biblical Chronologist 15.5 (November 11, 2025): 1–4. www.BiblicalChronologist.org.