|
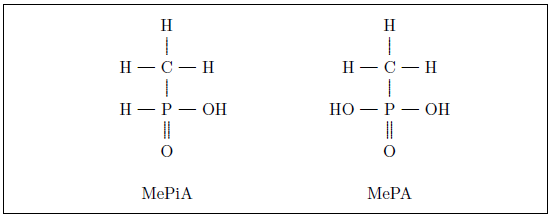
As discussed in Aging: Cause and Cure, methylphosphine gas (MeP) released into the atmosphere may be oxidized to methylphosphine oxide (MePO), which may be oxidized to methylphosphinic acid (MePiA), which may be oxidized to the stable end product methylphosphonic acid (MePA). As both MePiA and MePA are highly water soluble, they will leave the atmosphere together in rainfall, making them available to the human diet in drinking water from natural water sources such as rivers and lakes.
I was aware, when I first began to investigate atmospheric products of MeP back in 2014, that it was possible that either MePiA or MePA or both together might function in an anti-aging vitamin capacity. Ultimately, I launched a life span experiment with laboratory mice comparing MePA and MePiA at high daily intakes to help choose between these three options. At 24.7 weeks of age (0.48 years of age), a batch of 72, same age, female, ICR mice was randomly divided into two groups of 36 mice each and treatments were begun. One group was treated with 0.1 g MePA per liter in their drinking water and the other group was treated with 0.1 g MePiA per liter in their drinking water.
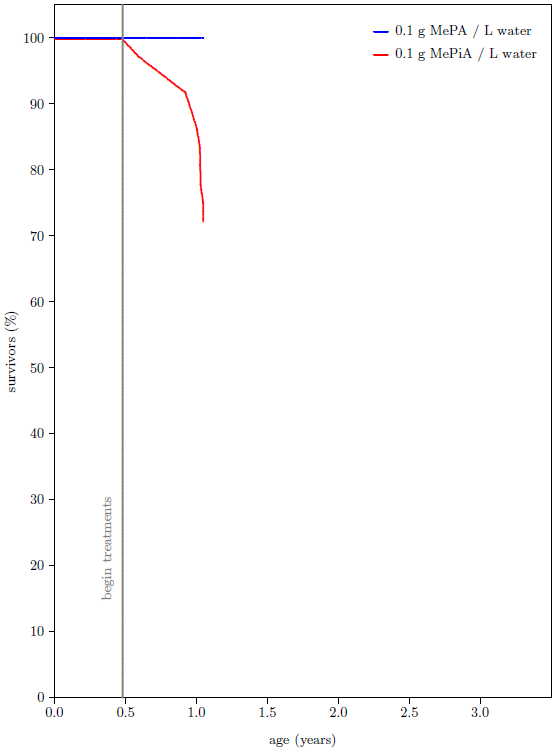
As I set about to write Aging: Cause and Cure, the state of this experiment looked as shown in Figure 2. The mice were just over a year old by that time, and they had been treated with MePA/MePiA for about six months. None of the MePA treated mice had died up to that point. In sharp contrast, more than a quarter of the MePiA treated mice had died. Relative to previous batches of similar mice raised over the course of nearly two decades in the ARP rodent lab, the MePA mice were doing unusually well and the MePiA mice were doing unusually poorly. It appeared, in fact, that MePiA was toxic to the mice. This seemed to settle the matter in favor of MePA being the sole anti-aging vitamin. Water-soluble vitamins are notoriously very low in toxicity. If MePiA was toxic at relatively low levels of intake, then it could not be a vitamin. This choice is embedded in Aging: Cause and Cure[2]
 |
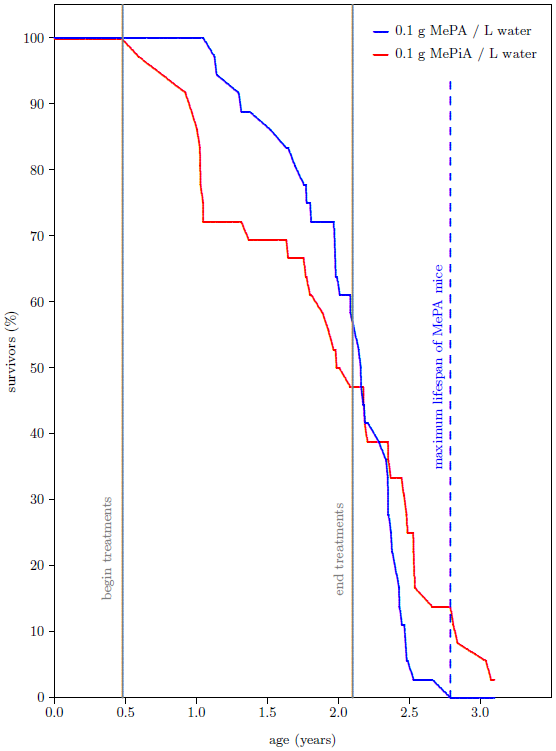
I kept the treatments going for another year, despite this negative result with MePiA. At 110 weeks (2.1 years) of age, roughly half of all of the mice had died. While the juvenile die-off of MePiA mice had ameliorated, MePA survival had consistently outperformed MePiA survival. There seemed no point in carrying the experiment further and the treatment regimen the mice had been on for over a year and a half was suspended. I expected all of the mice to be dead before another year had passed.
As a research technician was primarily responsible for mouse husbandry, I did not concern myself with these mice any further until a few weeks ago. At that point, with only three of the original 72 mice surviving, I happened to notice something which caught my attention. Two of the surviving mice looked to be very aged mice, as expected, but the third mouse looked younger than expected. This was not easy to explain. The best immediate idea was that someone had accidentally moved a younger mouse to the wrong cage at some point in the past. But, while this would ordinarily have been very hard to do, in this instance it had been simply impossible to do. Having concluded months previously that mice were unresponsive to what I then thought was the sole anti-aging vitamin, MePA,[3] I had decided to shut down the rodent lab. The batch of 72 mice used in this experiment had been the last batch of mice to enter the lab. They had entered the lab as weanlings. They were the youngest mice in the lab, and they were all of the same age. There had at no point been any younger mice available to accidentally put in the wrong cage. It appeared that this one mouse was, in fact, physiologically younger than her chronological age—that she had experienced anti-aging life lengthening.
The two elderly-looking mice have now both died. The younger-looking mouse is still alive. She is the last mouse in the rodent lab at present, and she is, in fact, the oldest mouse the lab has produced in its nearly two decades of operation.
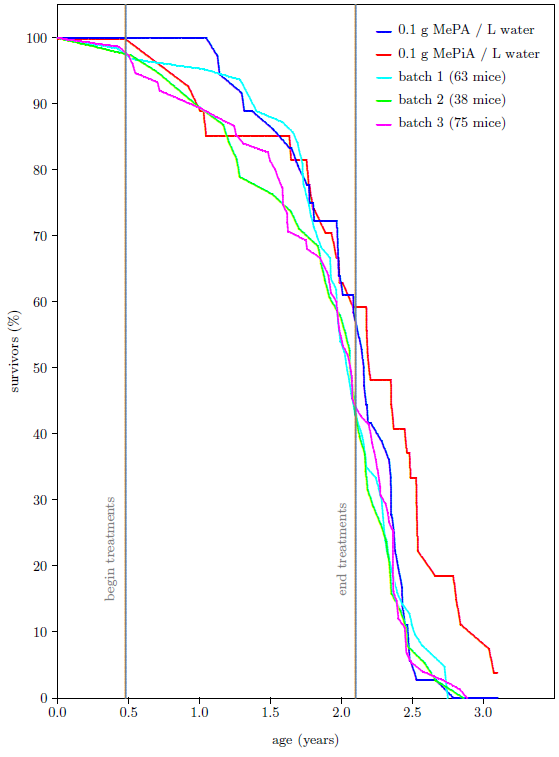
The survival curves for the 72 mice as they presently appear are shown in Figure 3. Contrary to expectation, the percentage of survivors in the group of MePiA treated mice ended up surpassing the percentage of survivors in the group of MePA treated mice. More importantly from the perspective of aging research,[4] the maximum life span for the MePiA mice is greater than the maximum life span for the MePA mice. Taken at face value, this shows life lengthening of the MePiA mice (including the two elderly-looking mice and the younger-looking mouse) relative to the MePA mice.
 |
Extension of maximum life span is extremely difficult to achieve. On the global research scene, only a very few interventions, such as calorie restricted diets, have succeeded in demonstrating an extension of maximum life span in mice. Though my aging research has attempted to demonstrate life lengthening for multiple decades with many hundreds of experimental animals—gerbils, fruit flies, vinegar worms, mice, and rats—this is the first time any evidence of increased maximum life span has been seen in my research. If it is real—if MePiA is indeed capable of extending the maximum life span of mice—then it is a major discovery.
Is this observed life-lengthening real, or is it just a statistical fluctuation? To answer this question, I added to the graph all of the data my lab has ever produced using batches of same age, weanling, female, ICR mice. There have been three additional batches in this category spanning nearly two decades. These batches had been subjected to a variety of treatments back in their day, intended to induce life lengthening (all of which had uniformly not succeeded in doing so). Figure 4 shows that the survival curve for the MePA mice of batch 4 is similar to the older three batches while the survival curve of the MePiA mice is clearly different. MePiA treatment does indeed appear to have extended the maximum life span for these mice.
This conclusion is at odds with the conclusion that MePiA is toxic to mice that was drawn two years ago from the juvenile portion of the survival curve. The toxicity conclusion obviously warrants further scrutiny.
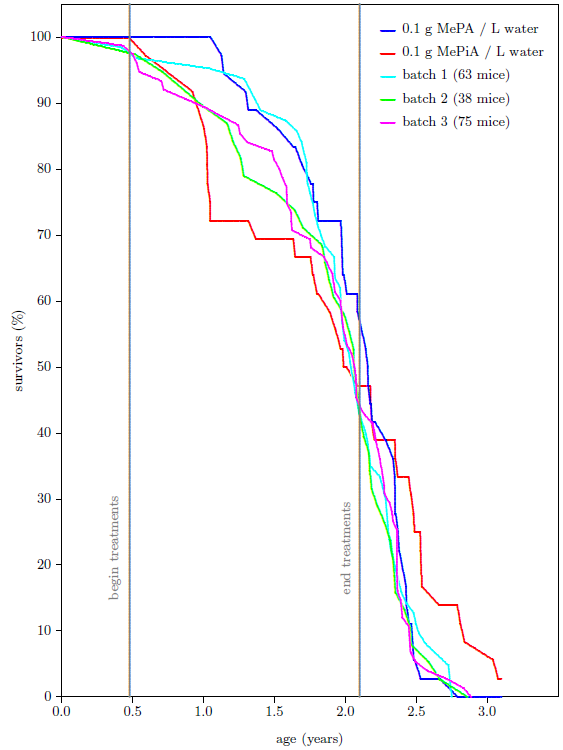 |
Upon detailed investigation of the early data, I found that the juvenile die-off seen in the data was dominated almost exclusively by a single cage of mice. The mice were housed in large cages, nine mice per cage. By the end of the juvenile die-off, only three mice remained in this one cage. The other three MePiA cages had six, eight, and nine survivors at this stage. This is not an impossible distribution of deaths due to toxicity, but it is improbable. In hindsight, the die-off seems more likely to have been a treatment-unrelated random event, possibly due to an infectious agent or to an overly-aggressive mouse in the depleted cage. This is supported by a subsequent, brief (14 weeks) treatment of another batch of mice (33 treated and 33 untreated) at ten times the dose (i.e., 1 g MePiA per liter) which gave no signs of toxicity. By the end of the 14 weeks, 1 treated mouse had died and 2 untreated mice had died.
When all mice from the depleted, infected(?) cage are excluded from the dataset, Figure 5 results. While both Figure 4 and Figure 5 show an extension of maximum life span for mice receiving MePiA, the Figure 5 graph probably displays the action of MePiA on these mice more accurately than the Figure 4 graph, so I will use it from this point on.
Now the question needs to be answered of why two of the three surviving, long-lived, MePiA treated mice appeared to be very elderly while the third appeared to be somewhat younger. Since these three mice were all on the same treatment, why did they not all show the same apparent physiological agedness?
 |
There are two possibilities. The first is that the two elderly-looking mice may have been dying of an unidentified wasting disease which gave them a particularly haggard appearance not due to physiological agedness. In support of this possibility is the fact that the two elderly-looking mice were raised together in the same cage, separate from the younger-looking mouse. Against this possibility is the fact that this elderly appearance has been characteristic of the relatively few long-lived, last-surviving mice I have observed in the ARP rodent lab, while it has not been seen with the relatively many mice dying at younger ages. This elderly appearance seems, in fact, to be uniquely diagnostic of advanced physiological age.
The second possibility is that, though all of these mice were of the same chronological age, the two elderly-looking mice were actually physiologically older than the younger-looking mouse. If this possibility is the true explanation, then two modes of action of the MePiA treatment are implied: a passive (diffusive) mode of action and an active (genetically programmed) mode of action for MePiA. The passive mode would always be in operation for all mice. The active mode would only be in operation if the genetics supporting it were intact.
In support of this second possibility is the fact that MePiA has not been environmentally available for the past 5500 years.[5] In consequence, there has been no selective pressure maintaining the genetically assisted mode for these thousands of years. Given the short generation time of mice, it is possible that the genetics needed for this mode to function might easily be missing or impaired for many modern mice. The more youthful appearance of the surviving mouse might result from this one mouse possessing MePiA-handling genetics (either by retention or by back-mutation) which the two elderly-looking mice did not possess.
If this second possibility is correct, then most of the life-lengthening displayed for mice older than two years in Figure 5 may be due to the passive mode of action. If so, then both the high treatment dose and the long duration of treatment which were used in this experiment were likely essential for obtaining the observed life-lengthening signal of the overall group. Meanwhile, the lone surviving mouse, in contrast, may have yielded an observable extension of maximum life span even at a significantly lower dose. A further implication in this second possibility is that this surviving mouse may go on living for many more weeks.
Regardless, the conclusion that supplementation with MePiA lengthens life spans of mice is robust. It does not rest on the observation of two distinct physiological states, nor does it rest on the ultimate fate of the surviving mouse. It rests on the empirical evidence of Figure 5 showing that a minimum of 50% of the MePiA mice lived longer than they would have without MePiA.
The postulate of passive and active modes of action, coupled with the ease with which MePiA may be oxidized, invokes Denham Harman's free radical theory of aging, in its final, mitochondrial form[6] as the fundamental explanation of the Figure 5 results.
Harman postulated that aging was due to accumulation of biomolecular damage due to free radicals (often called reactive oxygen species [ROS] in biological contexts) generated during normal cell metabolism. A strategy to limit damage due to free radicals is to make antioxidants (i.e., easily oxidized compounds) available to the cells. The antioxidants will then be on hand to "soak up" free radicals and thereby keep them from reacting destructively with cellular biomolecules.
Harman found that the average life span of mice could be increased by administering large daily intakes of select antioxidants to the mice. This corroborated the basic idea of free radical damage as a cause of aging. However, somewhat surprisingly, Harman found that maximum life spans of mice could not be increased in this way.
This led Harman to postulate that the mitochondria were the main site of life-limiting free radical damage and that possibly the mitochondria were inaccessible to his antioxidants.
Harman's theory now seems remarkably insightful and correct as far as it goes. It now appears that all that is missing is: 1. recognition that a particular antioxidant is genetically specified for the mitochondria, and 2. identification of that particular antioxidant.
MePiA, I suggest, is the genetically specified antioxidant required to complete Harman's theory.
This MePiA mitochondrial free radical theory of aging provides the following explanation of Figure 5.
For the first six months of their lives, before treatment with MePA / MePiA began, batch 4 mice aged at a normal rate due to mitochondrial degradation occasioned by free radicals generated within their mitochondria. Following initiation of treatments, MePA mice continued to age at a normal rate, MePA being resistant to oxidation and hence offering no protection against free radicals. Meanwhile, aging in MePiA mice was slowed due to the presence in their mitochondria of easily oxidized MePiA.
When treatment ceased, a normal rate of aging resumed in passive-mode MePiA mice. But the mitochondria of these MePiA mice had suffered less damage than the mitochondria of MePA mice throughout the treatment phase. As a result, the MePiA mitochondria were able to sustain post treatment damage for a longer time than the MePA mitochondria before critical damage levels, resulting in death, were reached. This evidenced as lengthened life spans of the MePiA mice relative to the MePA mice.
For any active-mode MePiA mice (possibly the younger-looking, surviving mouse), the outcome would have been even better. In this mode, the mitochondria are expected to possess the genetics necessary to actively support the natural antioxidant role played by MePiA in the mitochondria, maximizing MePiA's effectiveness against free radicals in that critical location. This includes the possibility of transport proteins able to actively transport MePiA molecules into the mitochondria, concentrating and keeping them there, as well as enzymes to recycle/reactivate MePiA molecules following their reaction with free radicals in the mitochondria. Thus, in the active-mode case, cessation of treatment does not result in immediate loss of mitochondrial protection. For active-mode MePiA mice, MePiA sequestered and recycled in the mitochondria may go on providing protection for a long time.
The MePiA mitochondrial free radical theory of aging provides an explanation not only of the Figure 5 data, but also of the human life span data mathematically modeled in Aging: Cause and Cure. Specifically, it explains the long biological half-life of the anti-aging vitamin found by that model.
The model finds a lifetime for MePA in the body of 194±4 years. The corresponding biological half-life is 135±3 years. This is a remarkably long biological half-life, which seems to imply specialized mechanisms for conservation of MePA and/or its metabolites.[7]
Updating "MePA" to "MePiA" everywhere in the above quote, one sees immediately that genetically programmed sequestering and recycling of MePiA in the mitochondria is precisely the sort of specialized conservation mechanism predicted by the model.
In addition, the passive and active modes of the MePiA mitochondrial free radical theory of aging explain why many of the MePiA mice of Figure 5 experienced so little life span gain from such large MePiA intakes.
The concentration of MePiA used in the mice experiment of Figure 5—100 mg/L—is expected to be on the order of 10,000 times larger than the concentration of MePiA naturally available in rainwater thousands of years ago. Meanwhile, the life spans of the treated mice were increased only about 20% at best, while the life spans of pre-Flood humans exceeded modern human life spans by at least 1,000%. Why have the mice yielded so little gain from so much treatment?
If it is assumed that the passive mode of MePiA action predominates with modern mice, for reasons explained above, while the active mode predominated with pre-Flood humans, then this difference in concentration may be explained as the difference in efficiencies of mitochondrial free radical scavenging between the passive and active modes.
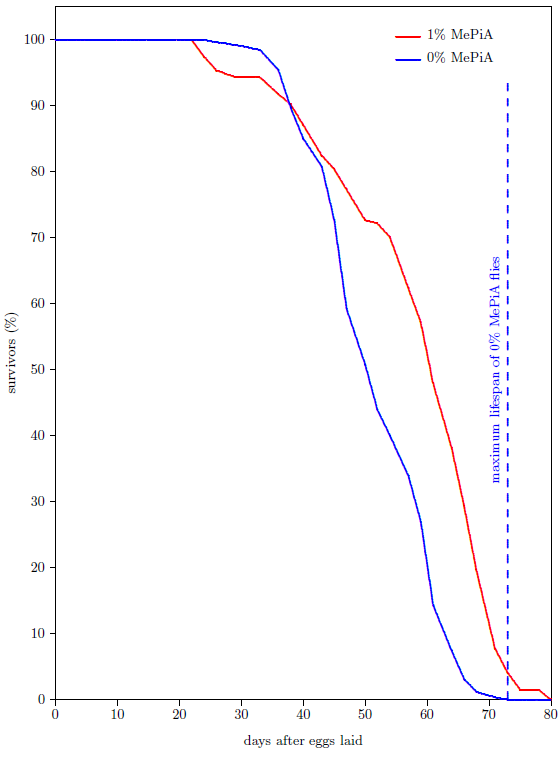 |
Because fruit flies are, like mice, eukaryotes possessing mitochondria, the MePiA mitochondrial free radical theory of aging predicts that fruit flies treated with MePiA will exhibit life lengthening. A life span experiment using fruit flies, shown in Figure 6, confirms this prediction.
In this experiment, the survival curve of 194 fruit flies raised together in a single cage on media containing 1% by weight of MePiA was compared with that of 260 fruit flies raised together in a single cage on media containing no MePiA. Not only was the treated flies' average life span increased, but, once again, extension of maximum life span was observed. Of the 194 MePiA flies, 8 were yet surviving when the last of the 260 untreated flies had died. The observed extension of maximum life span for MePiA treated flies in this case was 7 days.
While vitamin MePA has been found to bestow numerous health benefits on humans, the mice data of Figure 5 show that these benefits will not significantly increase maximum life span beyond that experienced with MePA deficient diets. To increase maximum life span, the diet must be supplemented with vitamin MePiA.
Because MePiA is easily oxidized to MePA, dietary supplementation with MePiA seems likely to make MePA also available by in vivo oxidation of MePiA to MePA. It may turn out, ultimately, that the body's entire need for MePA may be met in this way. For the immediate future, however, prudence dictates that both MePiA and MePA be supplemented together in the diet, as has been nature's custom from ancient times.
Inclusion of both MePA and MePiA in animal (including human) diets promises both to prevent and to cure aging as we know it and to greatly extend maximum life spans.
Yes it does. MePiA and MePA are indistinguishable to the model. This happens because they behave like twins in the environment. They both come from methylphosphine gas. They are both small, water-soluble acids. And they both leave the atmosphere in rainwater. Recall that Aging: Cause and Cure listed three vitamin X candidates: DMePiA, MePiA, and MePA.[8] All three were potentially able to satisfy the constraints of the model. MePiA was eliminated from consideration only because it was mistakenly thought to be toxic, based on the Figure 2 mice data.
No I am most certainly not saying that it doesn't do any good. It does lots of good. It just doesn't do, by itself, all the good which needs to be done to cure aging. If someone doesn't take the new MePA-plus-MePiA supplement because they want to use up their old MePA-only supplement first, they will be doing themselves relative harm. As soon as you have the new supplement, throw the old MePA-only supplement away.
No. I think that the discovery that MePiA is needed together with MePA is the complete answer, bringing to an end the quest to understand why pre-Flood humans were living so much longer than modern humans. Dietary supplementation to cure human aging as we now experience it is most unlikely to require anything other than vitamins MePiA and MePA.
No. In fact, the pace of research into these two anti-aging vitamins must now intensify. The fundamental discovery process has ended—the anti-aging vitamin duo has been discovered—but a great deal of "sweeping up" remains to be done. Most importantly, the difficult task of determining the optimum daily allowance must be tackled. At present, we are making our best estimate of this based on our total experience with these vitamins to date. But deliberate experiments are needed and are now underway to refine our knowledge and make sure these important vitamins are being used as effectively as possible to guarantee optimal health and maximal longevity. Changes to the recommended daily allowance and also to the relative concentrations of MePA and MePiA in the anti-aging dietary supplement seem likely outcomes of further research.